
 Pontos a serem abordados nesse artigo:
Pontos a serem abordados nesse artigo:1- Fertilidade de machos (ERCR)
2- Fertilidade de fêmeas (DPR)
3- Fórmula do novo 'net merit'
4- Estresse térmico
Introdução
Selecionar para fertilidade não é uma tarefa fácil, mas ao mesmo tempo é muito importante, pois reprodução, na maioria das fazendas, especialmente de gado leiteiro, é um dos principais motivos de preocupação. Para se ter uma idéia, um estudo em duas grandes fazendas no estado de São Paulo demonstrou índices de fertilidade baixos, com taxas de concepção em torno de 24% (no verão e outono) e 31% (no inverno e primavera) para vacas em lactação inseminadas artificialmente (Negrão et al., 2002). O objetivo desse artigo é fornecer as ferramentas necessárias para que um técnico possa fazer uso correto do grande volume de informações de fertilidade disponível hoje em dia. Para que isso ocorra, precisamos entender como são geradas as provas de fertilidade nos países exportadores de sêmen (vou utilizar os EUA como exemplo) e também estudar os trabalhos que avaliam a performance dos animais em situações de estresse térmico (maior obstáculo para fertilidade de vacas em várias regiões do Brasil).
Um aspecto importante com respeito às características de fertilidade é que elas são influenciadas por efeitos diretos (touro usado na inseminação) e efeitos maternos (influência do touro na capacidade de emprenhar de suas filhas). Nas avaliações genéticas, os efeitos diretos se referem à fertilidade de machos e os efeitos maternos à fertilidade de fêmeas.
Características como dias em aberto, intervalo entre partos e taxa de não-retorno ao cio têm baixa herdabilidade ( < 0,05), mas o coeficiente de variação das características de fertilidade é similar a de características de produção. O que gera essa baixa herdabilidade é o sistema de coleta de dados e de identificação dos animais para seleção (Royal et al., 2002). A taxa de não-retorno aos 70 dias se refere aos animais que após serem inseminados, não retornam a mostrar sinais de estro por 70 dias. Avaliando 1601 vacas holandesas, Vasconcelos et al. (1997) obtiveram ao redor de 20% de perdas embrionárias entre 28 e 98 dias de gestação, enquanto Fricke et al. (2003) reportaram 23% de perda de prenhezes entre 26 e 72 dias de gestação em 711 vacas holandesas checadas por ultra-sonografia, demonstrando a importância de se determinar quando as vacas foram diagnosticadas (70 dias de gestação, por exemplo).
O esforço de especialistas em reprodução em desenvolver esquemas de sincronização de cio e/ou ovulação, como por exemplo, o Ovsynch (Pursley et al., 1995), tem contribuído para a redução no intervalo entre partos, pois vacas são inseminadas mais cedo, mais frequentemente e dependendo menos da detecção de cio. Com o aumento constante da produção de leite por vaca e com a correlação positiva entre produção de leite e dias em aberto (+0,35), esta tarefa tende a se tornar mais difícil com o passar dos anos. A produção de leite por vaca praticamente dobrou nos EUA nos últimos 40 anos. Nesse período também foi verificado um aumento de 40 dias em aberto por lactação. Este aumento é parcialmente devido à seleção genética para produção de leite. Na figura 1, adaptada do trabalho de Washburn et al. (2002), mostra-se a tendência de aumento em produção de leite e de diminuição na taxa de concepção nas raças Jersey e Holandesa ao longo dos anos, além de ilustrar as diferenças e semelhanças entre essas raças com respeito a esses dois parâmetros.
A fertilidade de machos mede a habilidade do sêmen de um touro de gerar prenhezes, enquanto a fertilidade de fêmeas mede a habilidade das vacas em ciclar normalmente, mostrar sinais de estro e conceber através das várias lactações. Fertilidade de machos, fertilidade de fêmeas e manejo correspondem a aproximadamente 1%, 9% e 90% da variação em fertilidade, respectivamente, sendo que a nutrição e a detecção de cio são responsáveis por grande parte desses 90% e devem ser priorizadas se o objetivo principal for melhorar fertilidade.
1- Fertilidade de machos (ERCR)
A capacidade do espermatozóide de atingir o local de fertilização, fertilizar o ovócito e ativar o desenvolvimento embrionário são alguns dos pontos a serem medidos para se estimar precisamente a fertilidade de machos. As características relacionadas com fertilidade podem ser divididas em compensáveis e não-compensáveis. Características compensáveis (por exemplo motilidade e morfologia) são as que não afetam fertilidade se a quantidade de espermatozóides for aumentada na palheta de sêmen (Saacke et al., 2000). Essas características são fáceis de ser medidas, o custo dessas medições é baixo e os resultados são obtidos rapidamente. Devido aos altos padrões no que se refere à quantidade de espermatozóides por palheta de sêmen, na maioria das vezes, as características não-compensáveis são mais relacionadas com fertilidade do que as compensáveis. Maiores detalhes sobre características não-compensáveis podem ser encontrados na revisão de literatura feita por Braundmeier e Miller (2001), onde também são apresentadas as moléculas que afetam fertilidade, sua localização e função, porém, a maioria delas ainda não foi estudada em detalhe ou foi apenas estudada em humanos.
Determinar a fertilidade de machos através de ensaios laboratoriais é de valor limitado, porque o espermatozóide que vai fertilizar o ovócito é altamente selecionado, provavelmente não sendo representado pela média da amostra (sêmen) e muitas variáveis, algumas das quais nós não podemos controlar, interferem no processo de fertilização. Uma maneira melhor de se determinar a fertilidade de macho é a taxa de não-retorno aos 70 dias (quando se considera a vaca como prenhe). Nos EUA, a taxa de não-retorno aos 70 dias após a primeira inseminação está sendo utilizada através de um modelo animal (animal model), similar ao usado para se obter os PTAs de características de produção e tipo, para Holandesas e Jerseys (Dairy Records Management Systems - DRMS). Os dados analisados são ajustados para efeitos de idade da vaca, lactação, interação entre rebanho-ano-mês do parto, dias no leite, produção de leite corrigido para energia, nível genético da vaca e os resultados são publicados para todos os touros com mais de 300 inseminações (que já tem uma confiabilidade razoável) nos últimos 3 anos.
O "Estimated Relative Conception Rate" (ERCR) ou "Taxa de Concepção Relativa Estimada" pode ser encontrado na internet (www.drms.org). Esses resultados são publicados nas provas de Maio e Novembro e refletem a diferença, em porcentagem, de um touro com relação ao touro médio das companheiras de rebanho, com uma média de zero e cada unidade de ERCR corresponde a uma mudança de 1% na taxa de não-retorno. Os touros variam de -11% a +5% (16 pontos percentuais de diferença) nos resultados de 1998-2000. Comparados à média das primeiras inseminações das companheiras de rebanho (que foi considerado como 0), o touro mais fértil produziu um bezerro com 5% maior frequência e o touro menos fértil produziu um bezerro com 11% menor frequência. Cerca de 91% dos touros ficaram entre -3% a +3% e podem ser considerados como tendo fertilidade aceitável. Portanto, a ênfase de seleção dentro deste grupo não deve ser em fertilidade (www.drms.org).
Pecsok et al. (1994) concluíram que, nos EUA, um prêmio de 2 dólares (no valor da dose de sêmen) pode ser adicionado por unidade de acréscimo em ERCR, atingindo mais de 5 dolares em situações extremas. Na maioria das vezes, 2 touros devem se diferenciar por pelo menos 3 pontos em ERCR para serem considerados como tendo diferente ERCRs. Considerando que a maior parte da variabilidade em fertilidade é atribuída ao manejo e que outras características como NM$ (net merit) e PTA leite (habilidade predita de transmissão para leite) têm maior importância econômica, ERCR deve sempre ser considerado como uma característica secundária de seleção. Finalmente, pequenas diferenças em ERCR têm sido detectadas entre centrais de inseminação artificial, significando que o foco deve ser colocado em cada touro e não na central de inseminação que o comercializa. Weigel (2003) mostrou que a variação em médias de centrais é de 2% e a variação em cada central é de aproximadamente 10% (www.drms.org).
Com respeito a acurácia, os fatores determinantes são o número de inseminações e a repetibilidade, que por sua vez é derivada do número de inseminações e do desvio padrão estimado para ERCR. Por exemplo, existe uma probabilidade de 95% de que um touro com ERCR +1 e repetibilidade 60 ou 70 tenha o seu verdadeiro valor de ERCR entre -2 e +4. Touros com maior repetibilidade devem ser usados com maior intensidade do que touros de baixa repetibilidade.
Em um estudo feito nos EUA, utilizando 515.915 dados de inseminação de 517 rebanhos em 12 estados, Weigel (2003) mostrou que a vaca e o touro usado na inseminação foram responsáveis por 6% e 0,3% da variação fenotípica total em taxa de prenhez. Ao comparar inseminadores com pelo menos 1500 inseminações, a média variou de 15,8% a 41,1% e 2 inseminadores dentro de um mesmo rebanho variaram mais de 5% em alguns casos. A taxa de prenhez média em julho (verão nos EUA) foi de 25,9%, menor do que os 34,1% obtidos em fevereiro (inverno nos EUA). Vacas de alta produção também apresentaram menor fertilidade, vacas em primeira lactação com produção acima de 36 kg por dia e multíparas produzindo mais de 45 kg por dia tiveram 1,8% e 1,6% menor taxa de prenhez, quando comparadas a outros animais da mesma idade.
2- Fertilidade de fêmeas (DPR)
Em um estudo feito nos EUA, Weigel e Clay (2001) avaliaram 8,03 milhões de dados de inseminação de 1995 a 2001 e obtiveram uma variação de aproximadamente 10% em termos de PTA para fertilidade de filhas de touros holandeses, o que é muito próximo da variação obtida para fertilidade de machos e a correlação entre fertilidade de machos e fêmeas foi de 0,02. A correlação entre fertilidade de fêmeas e PTA leite foi de 0,09, o que significa que é possível selecionar para fertilidade de fêmeas, como uma característica secundária, sem grande perda em mérito genético para produção de leite.
A herdabilidade para fertilidade de fêmeas é de aproximadamente 4%, o que tem desestimulado a seleção direta para esta característica, apesar de que seleção indireta tem sido feita, nos EUA, através da vida produtiva (productive life). A correlação genética entre vida produtiva e dias em aberto é de aproximadamente -0,60, o que demonstra a tendência dos fazendeiros de descartar mais cedo animais que não emprenham.
A qualidade dos dados é uma grande preocupação em qualquer programa de seleção genética, sendo muito importante no caso de características de fertilidade, porque o julgamento errôneo de prenhez e as perdas embrionárias ocorrem rotineiramente no manejo de uma fazenda normal. Vacas que parem novamente podem ser facilmente checadas para dias em aberto, bastando subtrair 280 dias (prenhez média) do dia do parto.
Os resultados de "Daughter Pregnancy Rate" (DPR) ou taxa de prenhez de filhas, começaram a ser publicados nos EUA em fevereiro de 2003 e a unidade é 'dias em aberto'. O aumento de 1% em DPR corresponde a um aumento de 4 dias em aberto e vice-versa. Esta característica é influenciada pela capacidade das filhas de um touro de retornar a função reprodutiva normal após o parto, demonstrar sinais de estro e sua habilidade de conceber e manter a prenhez (Weigel e VanRaden, 2003). A tabela 1 mostra o aumento ou redução em dias em aberto (comparado com a média do rebanho no qual o sêmen vai ser usado) relacionado aos DPRs de vários touros importantes da raça holandesa. Os touros melhores diferem dos piores por 3 semanas de dias em aberto de suas filhas.
Touros jovens (com apenas uma geração de filhas) geralmente têm repetibilidades ao redor de 45% a 60%, com grande influência do pedigree nas suas provas. O PTA de touros para DPR varia mais do que outras características (produção de leite ou tipo, por exemplo), porque diversas fontes geram o PTA para DPR de um touro (inseminações relatadas, intervalo entre partos e descartes por reprodução) e o fato de que essas informações chegam em diferentes momentos durante a vida de um touro.
Provas de fertilidade geralmente são produzidas depois das provas de produção, devido ao intervalo necessário para que as filhas de um touro tenham oportunidade de emprenhar. Quando a informação de uma filha de um touro vem de um produtor que não reporta inseminações, o intervalo para essa informação ser incluída na prova (PTA para DPR) de um touro é ainda maior, porque essa vaca tem que parir para que se saiba quando ela emprenhou.
3- Fórmula do novo 'net merit'
A forte seleção para características de produção e sua correlação genética pouco favorável com fertilidade (dentro de rebanho), o fato de que rebanhos tendem a aumentar em tamanho (prejudicando a detecção de cio em alguns casos) e o fato de que as instalações hoje em dia obrigam as vacas a ficarem no concreto a maior parte do dia e as levam a mostrar menos sinais de estro, tem aumentado o enfoque dado às características de fertilidade. Nos EUA, a queda na taxa de concepção ao primeiro serviço (Washburn et al., 2002) e seu impacto na raça holandesa, principalmente nos últimos 20 anos, tem gerado grande preocupação, devido à grande importância desta raça. Os objetivos de seleção que eram restritos, na maior parte das vezes, à produção, tipo e algumas características de saúde, estão se modificando para incluir características de fertilidade, o que ocorre com o novo net merit (NM$), modificado em 2003.
A tabela 2 mostra os pesos dados para cada característica (www.aipl.arsusda.gov/reference/nmcalc.htm) e as novas características incluídas em 2003 no net merit (NM$). Essas modificações no net merit significam que seleção direta para fertilidade será realizada através da taxa de prenhez de filhas (daughter pregnancy rate), que recebe 7% do peso total do índice. Esta seleção vinha sendo feita de maneira indireta, através da vida produtiva (productive life), nos EUA e nos países que usam os EUA como uma das fontes de material genético (o que é o caso do Brasil). Touros são ranqueados pelo net merit e este índice também influencia o preço do sêmen. A correlação genética entre as provas de 2000 e 2003 é de 0.98 (www.aipl.arsusda.gov/reference/changes/eval0308.html).
As habilidades preditas de transmissão (PTA) do novo net merit, cheese merit e fluid merit têm uma correlação de 0,15, 0,17 e 0,12 com taxa de prenhez de filhas, o que gera um aumento esperado em PTA de 0,05 por ano e uma mudança em valor genético de 1,0 por década (www.aipl.arsuda.gov). A mudança esperada em valor genético é igual ao ganho esperado em PTA multiplicado por dois e neste caso multiplicado por 10 para expressar o valor em décadas.
4- Estresse térmico
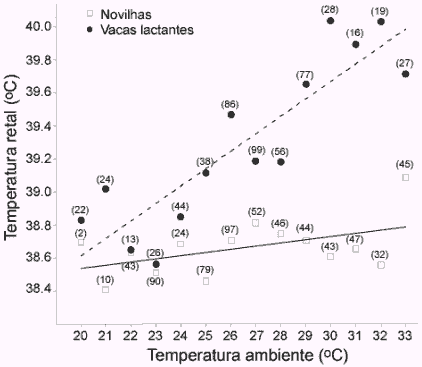
O estresse térmico é motivo de grande preocupação em quase todas as regiões do Brasil, levando animais a renderem menos em reprodução e produção. Nessas condições, vacas reduzem ingestão de alimentos e atividade física, mostrando menos sinais de estro. Estresse calórico ocorre, especialmente em vacas em lactação (veja figura 2; Sartori et al., 2002) quando animais não são capazes de manter sua temperatura corporal quando a temperatura ambiente aumenta. Esse aumento em temperatura corporal acaba afetando diversas funções corpóreas. Diversos estudos têm mostrado que o aumento de temperatura no momento da inseminação reduz fertilidade e aumenta a perda embrionária, a viabilidade dos ovócitos e dos espermatozóides é prejudicada, embriões jovens são afetados (principalmente nos primeiros dias de vida), a aceitação de monta é reduzida e a intensidade e duração do estro também. Em um estudo da Universidade de Wisconsin-Madison, Sartori et al. (2002) observaram uma taxa de fertilização de apenas 55% em vacas holandesas lactantes durante o verão e de aproximadamente 90% durante o inverno.
Vacas sob estresse térmico, apresentam redução nos níveis sangüíneos de estradiol antes do cio, redução no tamanho do folículo dominante da segunda onda folicular, maior número de ondas foliculares por ciclo estral e fases luteais mais longas (Wilson et al., 1998). Ravagnolo e Misztal (2003) avaliando 18.059 taxas de não-retorno aos 45, 60 e 90 dias após a primeira inseminação e 81.674 dados de produção de leite (test-day) na Flórida, notaram que a taxa de não-retorno aos 90 dias (NR90) gera a maior herdabilidade e que sua correlação genética com produção de leite foi de -0,41. Essa relação antagônica significa que é possível se realizar seleção para ambas as características simultaneamente. A tolerância ao calor para NR90 e a tolerância ao calor para produção de leite são provavelmente diferentes processos metabólicos, porque sua correlação genética foi de 0,04. Ravagnolo et al. (2000) mostraram que as taxas de não-retorno foram menores quando o índice de temperatura e humidade (THI) ultrapassou 66 ou 70 (dependendo do estado nos EUA) e que cada ponto de aumento em THI, após esses limites, gerou uma redução de 0,005 e 0,007 em taxas de não-retorno {THI(t,h)=9/5t+32-11/2(1-h)(9/5t-26)(NOAA, 1976)}.
Ravagnolo e Misztal (2003), avaliando 150.200 dados de primeira inseminação de 110.860 vacas holandesas de 550 rebanhos na Geórgia (EUA), mostraram que o THI do dia da inseminação teve o maior efeito em NR45, seguido de 2 dias antes, 5 dias antes e 5 dias após a inseminação, mas nenhuma correlação foi encontrada 10, 20 e 30 dias após a inseminação. Animais em sua primeira lactação foram mais susceptíveis ao aumento em THI do que os animais em lactações mais avançadas (0,008 vs 0,005 decréscimo para cada ponto de aumento em THI). Esse estudo também mostrou que animais com 150 dias de lactação tiveram 0,16 menos NR45 do que animais com menos de 60 dias em lactação na sua primeira inseminação. Ventiladores e aspersores de água são alguns dos métodos utilizados para combater o estresse térmico, mas eles geralmente não eliminam o problema por completo e têm custo alto de implementação.
Tabela 1. Exemplos de touros holandeses, seu DPR e o correspondente valor em dias em aberto.
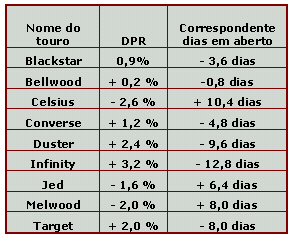
Tabela 2. Pesos dados para cada característica no cálculo do velho (2000) e do novo (2003) net merit.

Figura 1. Produção de leite e taxa de concepção em 532 rebanhos holandeses e 29 rebanhos Jersey de 1976 a 1999 (Washburn, 2002).
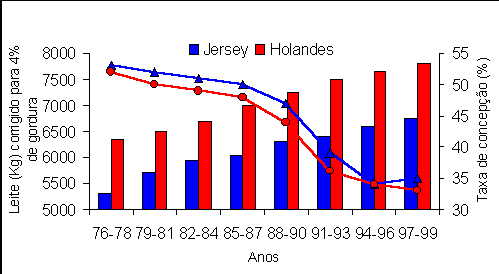
Figura 2. Relação entre temperatura ambiente e temperatura corporal em vacas lactantes e novilhas nulíparas da raça Holandesa (Sartori et al., 2002). Os dados entre parênteses representam o número de observações por ponto.
Referências
A. G. Braundmeier, D. J. Miller. The Search is on: Finding Accurate Molecular Markers
of Male Fertility. J. Dairy Sci. 84:1915-1925,2001.
J. L. M. Vasconcelos, R. W. Silcox, J. A. Lacerda, J. R. Pursley, M. C. Wiltbank. Pregnancy rate, pregnancy loss, and response to heat stress after AI at 2 different times from ovulation in dairy cows. Biol. Reprod. 56 (Supl. 1):140, 1997.
J. R. Pursley, M. O. Mee, M. C. Wiltbank. Synchronization of ovulation in dairy cows using PGF2? and GnRH. Theriogenology 44:915-923, 1995.
K. A. Weigel, J. S. Clay. Possibilities for genetic improvement of fertility in US dairy cattle. J. Dairy Sci. 84 (Suppl. 1):245, 2001.
K. A. Weigel, P. VanRaden. New Evaluations Offer a Genetic Approach to Improving Cow Fertility (www.naab-css.org), 2003.
K. A. Weigel. Improving the reproductive efficiency of dairy cattle through genetic selection. J. Dairy Sci., 2003 (enviado para publicação).
M. D. Royal, J. E. Pryce, J. A. Woolliams, A. P. F. Flint. The Genetic Relationship between Commencement of Luteal Activity and Calving Interval, Body Condition Score, Production, and Linear Type Traits in Holstein-Friesian Dairy Cattle. J. Dairy Sci. 85:3071-3080, 2002.
M. D. Royal, A. P. Flint, J. A. Woolliams. Genetic and phenotypic relationships among endocrine and traditional fertility traits and production traits in Holstein-Friesian dairy cows J. Dairy Sci. 85:958-967, 2002.
O. Ravagnolo, I. Misztal. Effect of Heat Stress on Nonreturn Rate in Holsteins: Fixed-Model Analyses. J. Dairy Sci. 85:3101-3106, 2002.
O. Ravagnolo, I. Misztal. Effect of Heat Stress On Nonreturn Rate in Holstein Cows: Genetic Analyses. J. Dairy Sci. 85:3092-3100, 2002.
O. Ravagnolo, I. Misztal, G. Hoogenboom. Genetic Component of Heat Stress in Dairy Cattle, Development of Heat Index Function. J. Dairy Sci. 83:2120-2125, 2000.
P. M. Fricke, D. Z. Caraviello, K. A. Weigel, M. L. Welle. Use of Ovsynch for resynchronizing ovulation at various intervals after first postpartum timed artificial insemination in lactating dairy cows. J. Dairy Sci., 2003 (enviado para publicação).
R. Sartori, R. Sartor-Bergfelt, S. A. Mertens, J. N. Guenther, J. J. Parrish, M. C. Wiltbank. Fertilization and early embryonic development in heifers and lactating cows in summer and lactating and dry cows in winter. J. Dairy Sci. 85:2803-2812, 2002.
S. J. Wilson, C. J. Kirby, A. T. Koenigsfeld, D. H. Keisler, M. C. Lucy. Effects of controlled heat stress on ovarian function of dairy cattle. 2. Heifers. J. Dairy Sci. 81:2132-2138, 1998.
S. J. Wilson, R. S. Marion, J. N. Spain, D. E. Spiers, D. H. Keisler, M. C. Lucy. Effects of controlled heat stress on ovarian function of dairy cattle. 1. Lactating cows. J. Dairy Sci. 81:2124-2131, 1998.
S. L. Negrão, J. R. Chiari, D. G. B. Demétrio, C. A. Rodrigues. Therapeutic embryo in high production dairy cows with more than three artificial inseminations. Theriogenology 57:555, 2002.
S. P. Washburn, W. J. Silvia, C. H. Brown, B. T. McDaniel, A. J. McAllister. Trends in reproductive performance in southeastern Holstein and Jersey DHI herds. J. Dairy Sci. 85:244-251, 2002.
S. R. Pecsok, M. L. McGilliard, R. L. Nebel. Conception rates. 2. Economic value of unit differences in percentages of sire conception rates. J. Dairy Sci. 77:3016-3021, 1994.

