
 FAÇA SEU LOGIN E ACESSE CONTEÚDOS EXCLUSIVOS
FAÇA SEU LOGIN E ACESSE CONTEÚDOS EXCLUSIVOS
Introdução – Comportamento de estro
O estro representa o período de receptividade sexual, que é seguido por ovulação. A expressão do estro é caracterizada por uma série de comportamentos, alguns dos quais são mais específicos que outros. Como a ovulação em vacas ocorre cerca de 30 horas depois da manifestação dos sinais de estro (Roelofs et al., 2005), a capacidade de reconhecer estes comportamentos e detectar o estro é crítica para a determinação do momento da IA. A detecção do cio é um componente essencial dos programas reprodutivos e as baixas taxas de detecção já são uma preocupação antiga (e.g. Helmer e Britt, 1985).
Principal comportamento
O estradiol é o hormônio indutor do comportamento estral: em vacas, o estradiol 17-β é o hormônio responsável pela indução do comportamento estral e ovulação. Estudos conduzidos com vacas ovariectomizadas e intactas demonstraram que as injeções de várias dosagens de estradiol induziam a expressão de comportamento em intensidades semelhantes (Allrich, 1994), o que permitiu concluir que o efeito do estradiol sobre o comportamento estral ocorre de maneira “tudo ou nada”: o comportamento estral é induzido uma vez que o estradiol atinge uma determinada concentração fixa (limiar) na circulação sistêmica. O aumento da concentração acima deste limiar, o que não foi quantificado, não altera a expressão de comportamento estral (Allrich, 1994).
O principal sinal de estro em vacas é a imobilidade quando montadas por outras fêmeas (Roelofs et al., 2010), também denominado reflexo de imobilidade (Albright e Arave, 1997). Pesquisas das décadas de 70 e 80 relataram maior frequência e maior duração de observação visual de imobilidade para a monta que os níveis observados atualmente (Figura 1 a, b, c). Embora pareça óbvio que a manifestação da imobilidade e aceitação da monta sejam menos frequentes hoje em dia, não devem ser consideradas as únicas manifestações a serem avaliadas para julgar a intensidade de estro. O comportamento estral varia com o número de animais em estro, qualidade do piso e tamanho do grupo de vacas (Albright e Arave, 1997). A atividade de monta, por exemplo, é muito mais frequente em piquetes abertos ou espaços com piso de terra (Britt et al., 1986), com cerca de 80% da atividade de monta observada no curral de vacas secas (Pennington et al., 1985).
Figura 1 - a) Frequência de imobilidade (aceitação da monta) por estro, b) duração da imobilidade (h), e c) frequência de monta por estro segundo relatos de 1975 a 2011
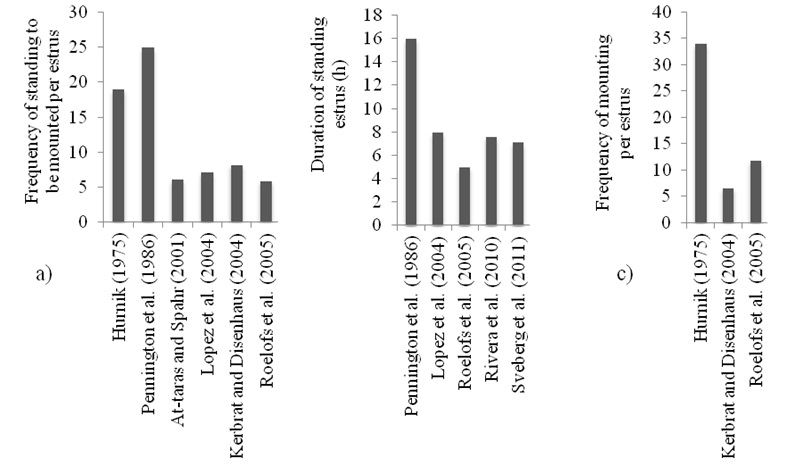
Ao longo dos últimos 30 anos, a observação da atividade de monta revela mudanças em manifestações comportamentais do estro, provavelmente devido à qualidade das instalações e maior confinamento das vacas. É claro que as vacas leiteiras modernas, considerando as atuais condições das fazendas leiteiras, não aceitam a monta com a frequência que seria necessária para a avaliação puramente visual do estro. As alternativas a serem consideradas são: 1) oferecer às vacas um ambiente que estimule a atividade de monta, 2) melhorar/ e desenvolver métodos para a detecção das montas e 3) avaliar métodos de detecção de comportamentos alternativos. Estas duas últimas alternativas receberam bastante atenção nos últimos 10 anos, com o uso de detectores eletrônicos de monta e monitores automáticos de atividade (MAA).
Cronologia dos eventos fisiológicos relacionados ao estro
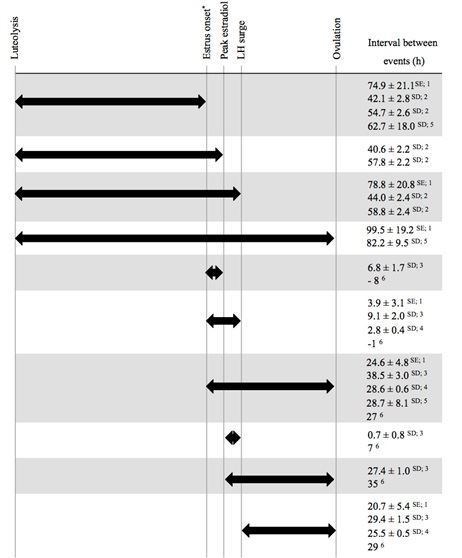
O tempo entre luteólise e ovulação é regulado (Figura 2) por diversos fatores, o que leva à alta variabilidade destes intervalos e pode ser uma das causas de baixas taxas de concepção (Saumande e Humblot, 2005). Supondo um intervalo de 28 horas entre o estro e a ovulação e 25 horas entre o pico de LH e a ovulação (Bloch et al., 2006), pode-se concluir que o pico de estradiol, pico de GnRH e pico de LH ocorrem em um período de apenas 3 horas. No momento em que termina o comportamento de cio, a ovulação é o único evento que ainda vai ocorrer para completar o ciclo. O início do período de maior atividade (medido por MAA) ocorre 29 horas antes da ovulação (Valenza et al., 2012). Uma fazenda que utiliza MAA insemina as vacas 7 a 12 horas depois do início do pico de atividade (Neves e LeBlanc, 2015), deixando um intervalo de cerca de 10 horas entre a IA e a ovulação.
A grande variação quanto ao momento de ocorrência da ovulação em relação à manifestação do cio poderia ser a explicação para as baixas taxas de concepção (Valenza et al., 2012). Por sua vez, a grande variação na duração do estro e momento de manifestação de atividade de pico neste período poderia também contribuir para a variabilidade no momento da ovulação. Usando um sensor diferente, Stevenson et al. (2014) relataram intervalo semelhante entre manifestação do estro e ovulação (26 horas), mas com menor variação. Interessante observar que, enquanto Valenza et al. (2012) avaliaram o estro depois de tratamento hormonal, Stevenson et al. (2014) estudaram o estro espontâneo. Seria de se esperar que a sincronização do estro reduzisse a variabilidade do tempo decorrido entre eventos endócrinos e fisiológicos, mas os tempos foram muito semelhantes. Stevenson et al. (2014) também relataram tempos semelhantes entre a manifestação do estro e a ovulação quando o estro foi determinado pelo início de aumento de atividade ou primeiro reflexo de imobilidade e aceitação da monta (24,6 ± 0,7 horas vs. 26,4 ± 0,7 horas, respectivamente).
O pico de LH interrompe a atividade da aromatase e interrompe a síntese de estradiol pelo folículo (Forde et al., 2011). Da mesma forma, observa-se uma redução de 50% das concentrações de estradiol em relação ao pico às 5 horas pós-pico de LH (Chenault et al., 1975). Às 14 horas pós-pico de LH, o estradiol já está praticamente em níveis basais (2 pg/mL; Chenault et al., 1975) e abaixo do limiar de 2 desvios padrões acima da linha basal (Aungier et al., 2015).
Em relação ao estro comportamental, foi observado que a concentração de estradiol atinge o pico no momento da máxima expressão comportamental de cio (Van Eerdenburg et al., 1996), mas cai em seguida para 60% dos valores de pico em 6 horas após máxima expressão de cio (Lyimo et al., 2000). Pode-se extrapolar que as últimas horas de estro comportamental ocorrem em face de concentrações decrescentes de estradiol circulante. Entretanto, não está claro por que não existe correlação linear entre a intensidade do estro e concentrações de estradiol (Aungier et al., 2015; Madureira et al., 2013; Silper et al., 2015c).
Papel do estradiol e da progesterona
A elevação da concentração circulante de estradiol suprarregula a expressão de receptores de progesterona no endométrio ao estabilizar o mRNA do receptor α de estradiol (ERα) (Ing e Ott, 1999). A liberação constante de progesterona por um período relativamente longo (10-14 d) durante o diestro induz a redução da expressão gênica de ambos os receptores (Spencer e Bazer, 1995). De maneira geral, a dinâmica de hormônio e receptor, tanto para a progesterona quanto para o estradiol, é essencial para criar um ambiente ideal no endométrio e uma gestação bem-sucedida.
A concentração circulante de estradiol não está relacionada de forma linear com o comportamento estral e é provável que outros fatores contribuam para a variação observada no comportamento estral. Ainda que a indução de comportamento estral por injeção de estradiol em diferentes dosagens tenha resultado em expressão comportamental semelhante do estro (Allrich, 1994), não se pode supor que a quantidade de estradiol que atua no hipotálamo tenha sido a mesma em todas as vacas que receberam a mesma dosagem. Variação comportamental individual, taxa metabólica, esteroides endógenos, desenvolvimento neuronal e expressão de receptores de estradiol são algumas das potenciais fontes de variação na expressão do estro, além das concentrações circulantes de estradiol. Além disso, é possível que a menor dosagem testada de estradiol já tenha sido suficiente para induzir um grau satisfatório de expressão comportamental.
Nos bovinos, somente a distribuição de ERα foi estudada. Os receptores ERα são expressos nas mesmas áreas do hipotálamo descritas em outras espécies, durante a fase lútea (van Eerdenburg et al., 2000). Durante estro e meta-estro, entretanto, a expressão de ERα foi detectada somente no núcleo arqueado. Segundo os autores, a ausência de expressão de ERα no estro e em vacas ovariectomizadas exclui a possibilidade de auto infrarregulação dos receptores e pode indicar por que a expressão de estro é reduzida em vacas leiteiras. A ausência de coloração para ERα também foi relacionada à possibilidade de que os receptores estavam ocupados pelo ligante, mas esta não foi considerada uma hipótese robusta. Coletivamente, van Eerdenburg et al. (2000) concluíram que estradiol e ERα são fatores-chave na sincronização de eventos reprodutivos fisiológicos e comportamentais. O controle do comportamento estral depende da expressão de ERα em núcleos hipotalâmicos específicos, assim como de concentrações de estradiol altas o suficiente para desencadear reflexos de comportamento sexual.
O papel da progesterona no priming do cérebro bovino para as ações do estradiol é evidenciado pela ausência ou baixa expressão do estro na primeira ovulação pós-parto e no primeiro estro da puberdade, eventos que não são precedidos por uma fase lútea. Expressão mais intensa do estro com protocolos de IATF que incluam dispositivos vaginais de progesterona (CIDR) confirma o papel da progesterona como um primer para a indução do comportamento estral desencadeado pelo estradiol (Rhodes et al., 2002). Já foi criada a hipótese de que, em cada ciclo, as concentrações de estradiol durante o estro iriam induzir um estado cerebral refratário, que seria revertido pelo pico de progesterona da fase lútea (Allrich, 1994; Woelders et al., 2014). Estradiol e progesterona induzem comportamento estral em vacas leiteiras ovariectomizadas secas (Vailes et al., 1992). Entretanto, os autores observaram que a intensidade do estro comportamental é mais complexa e vai além da relação entre estradiol e progesterona.
A indução de um perfil de altos níveis de progesterona e estradiol reduz mais a imobilidade que a monta, em comparação a vacas somente sob ação do estradiol (Vailes et al., 1992). Provavelmente, o estradiol não é o único fator a regular a expressão do estro (Roelofs et al., 2010), mas é necessário para desencadear o processo. Ainda que altas concentrações de estradiol não intensifiquem a expressão do estro, é provável que melhorem a função do trato reprodutivo (Allrich, 1994).









