
 FAÇA SEU LOGIN E ACESSE CONTEÚDOS EXCLUSIVOS
FAÇA SEU LOGIN E ACESSE CONTEÚDOS EXCLUSIVOS


Introdução
No mundo todo, a perda embrionária pode ser responsável pela maior perda econômica isolada para os produtores de bovinos. Com 40 milhões de vacas e novilhas de corte expostas anualmente à reprodução nos Estados Unidos, as perdas anuais devidas à mortalidade embrionária são superiores a 2,2 bilhões de reais. No Brasil, com mais de 70 milhões de vacas de corte anualmente em reprodução, as perdas causadas pela mortalidade embrionária podem ser duas vezes maiores.
Os mecanismos envolvidos no estabelecimento e manutenção da prenhez são complexos e, tomando por base a literatura, foram poucos os progressos alcançados nos últimos 90 anos para reduzir o desperdício de embriões. O setor de produção leiteira dos Estados Unidos está testemunhando um aumento na perda embrionária. Para evitar os problemas de fertilidade que a indústria leiteira está enfrentando, os pecuaristas de corte precisam estar mais atentos a estes problemas e tornar-se proativos para preveni-los. Este trabalho irá focar as causas e a época em que ocorrem as falhas de prenhez e discutir algumas práticas de manejo que poderiam reduzir estas perdas.
Em geral, é aceito que as taxas de fertilização em bovinos depois de uma única cobertura estejam entre 90 e 100%, quer seja usada a monta natural ou a inseminação artificial (Sreenan e Diskin, 1983). Entretanto, é raro que 70% das coberturas individuais resultem em diagnóstico positivo de prenhez 30 dias mais tarde e um número ainda menor resulte em um bezerro nascido vivo. As perdas embrionárias são definidas como as perdas que ocorrem entre a fertilização até o dia 42 da prenhez, quando a diferenciação e a implantação já ocorreram. As perdas que acontecem após o dia 42 são geralmente denominadas perdas fetais.
As perdas embrionárias são ainda divididas em duas categorias e classificadas como Mortalidade Embrionária Precoce (MEP; da fertilização até o dia 27) e Mortalidade Embrionária Tardia (MET; do dia 28 ao 42). A maior parte da mortalidade embrionária é representada pela mortalidade embrionária precoce, com relato de taxas variando de 20 a 40% em bovinos de corte (Humbolt, 2001). A mortalidade embrionária tardia ocorre em 3 a 14% de vacas e novilhas de corte (Humbolt, 2001; Perry et al., 2005; dados não publicados). Existe grande quantidade de informações sobre a mortalidade embrionária em bovinos de leite, mas há muito poucas informações disponíveis em bovinos de corte. Maurer e Chenault (1983) sugerem que a falha na gestação é de tipo diferente quando comparamos vacas e novilhas de corte. Neste estudo, a maior parte das falhas na gestação em novilhas foi devida a falha na fertilização mais do que a perda embrionária. A taxa de recuperação de embrião ou de óvulo não fertilizado foi mais baixa em novilhas do que em vacas, sugerindo que talvez uma parte das novilhas não ovulou depois de estarem no cio. A medida precisa da mortalidade embrionária é complicada ainda pelo fato de que não conseguimos determinar o diagnóstico da prenhez antes do dia 27, aproximadamente (por ultra-som ou indicadores específicos de prenhez presentes no sangue), sem fazer a coleta do trato reprodutivo ao abate.
Não há nenhum fator isolado que tenha sido comprovado como capaz de prevenir a mortalidade embrionária precoce. Entretanto, se pudermos prevenir a perda do embrião em 5 de cada 100 vacas, poderíamos desmamar um adicional de 950 kg.
Entendendo o ciclo estral e o início da gestação
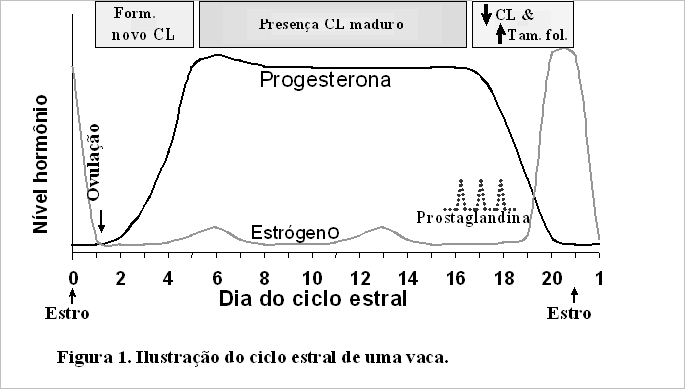
Uma revisão do ciclo estral (Figura 1) e do início da prenhez (Figura 2) de uma vaca podem ser úteis para entender algumas das causas de perda embrionária. No dia 0 ou 21 do ciclo estral, a fêmea está no cio e um de seus ovários tem uma estrutura semelhante a uma bolha que recebe o nome de folículo, e que contém um óvulo maduro. Este folículo está produzindo altos níveis de estrógeno, fazendo com que a vaca apresente sinais comportamentais de cio. No dia 1, aproximadamente 28 horas após o início do cio, este folículo dominante se rompe. O óvulo é liberado e penetra no oviduto, onde a fertilização pode ocorrer se houver a presença de espermatozóides viáveis.
Se a fertilização não ocorrer, o óvulo não fertilizado irá percorrer o oviduto, chegando ao útero em torno do dia 7. Com a liberação do óvulo do ovário, as células do folículo rompido sofrem uma reorganização (luteinização) e tornam-se células luteais que crescem e se dividem ao longo dos 5 dias seguintes, preenchendo a cavidade folicular no ovário, para formar uma estrutura denominada CL (corpo lúteo). O CL produz progesterona, que prepara o útero para uma gestação.
Gradativamente, os níveis de progesterona se elevam do dia 1 ao dia 5, inibindo a liberação do hormônio liberador de gonadotrofina (GnRH), do hormônio luteinizante (LH) e do hormônio folículo estimulante (FSH) em quantidades suficientes para permitir a maturação final dos novos folículos nos ovários. Assim, estes folículos recém formados tornam-se atrésicos. Ao longo dos 12 dias seguintes, o CL maduro produz progesterona e os níveis de progesterona no sangue permanecem elevados, prevenindo a maturação final dos folículos em desenvolvimento.
Quando não há embrião presente no útero aproximadamente no dia 17 do ciclo estral, o útero produz o hormônio prostaglandina, que destrói o CL, diminuindo os níveis de progesterona (Figura 1). A baixa progesterona permite que os pulsos de GnRH, FSH e LH aumentem, proporcionando o suporte hormonal para o desenvolvimento dos folículos nos ovários, um folículo torna-se dominante (folículo pré-ovulatório) e produz estrógeno, resultando em nova ocorrência de estro e ovulação.
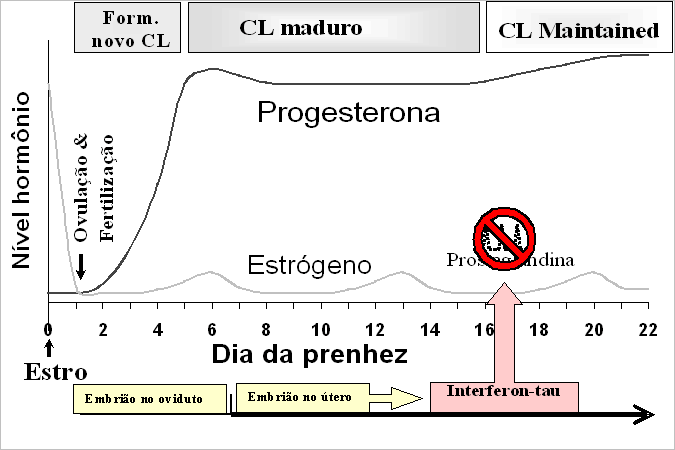
Se ocorrer a fertilização, o embrião fertilizado irá dividir-se e percorrer o oviduto, alcançando o útero no dia 7, quando se espera que o embrião seja implantado e desenvolva-se até chegar a bezerro. Para sobreviver, o embrião precisa continuar a dividir-se, alongar e produzir interferon-tau que bloqueia a produção uterina de prostaglandina, em um processo denominado reconhecimento materno da prenhez (Figura 2). Em geral, os embriões produzem interferon-tau do dia 14 ao dia 23, mas precisam produzir o suficiente para pelo menos prevenir a liberação uterina de prostaglandina e a destruição do corpo lúteo em torno do dia 17 a 18 depois do estro.
Qualquer coisa que impeça o crescimento do embrião ou a produção de interferon-tau ou qualquer coisa que interfira com a capacidade uterina de reconhecer este sinal ou bloquear a prostaglandina, poderia impedir o reconhecimento materno da prenhez e resultar em retorno ao estro, ainda que um embrião esteja presente. Se não ocorrer o reconhecimento materno da prenhez, o embrião irá morrer e a vaca retornará ao cio, geralmente sem qualquer atraso na duração do ciclo estral.
Figura 2. Ilustração da fase inicial da prenhez em uma vaca.
Fatores genéticos que afetam a perda embrionária
As anomalias genéticas respondem por aproximadamente 10% das perdas embrionárias e geralmente resultam em falha na prenhez nas duas primeiras semanas (King et al, 1990). A expressão de genes letais pode causar a morte do embrião nos primeiros 5 dias da gestação. Um outro transtorno genético que contribui para a morte embrionária é um número anormal de cromossomos em algumas ou em todas as células embrionárias. Esta é a anomalia genética mais comum e geralmente resulta na morte embrionária.
As causas do número anormal de cromossomos incluem polispermia (quando mais de um espermatozóide fertiliza um óvulo) e erros meióticos nos gametas ou no embrião em desenvolvimento. Pode haver uma incidência mais elevada de polispermia quando as inseminações artificiais ocorrem mais próximas do momento da ovulação, não dentro do período ideal de 12 horas após o início do estro (Saacke et al., 2000).
Em geral, os espermatozóides precisam de cerca de 6 horas no trato reprodutivo da fêmea para que ocorra a capacitação, antes que tenham capacidade de fertilização. Assim sendo, quando as inseminações ocorrem próximo do momento da ovulação, o ovócito está disponível para a fertilização, mas precisa esperar que ocorra a capacitação do espermatozóide. Durante este período de espera, o ovócito "envelhece" e começa a perder sua capacidade de bloquear a polispermia. Mesmo que o ovócito seja capaz de fazer o bloqueio, este envelhecimento parece resultar em um atraso ou interferência no desenvolvimento do embrião e, desta forma, diminui a qualidade do embrião.
Na verdade, a inseminação realizada mais próxima do momento da ovulação resulta em taxa de fertilização mais elevada, mas em qualidade embrionária menor. As inseminações mais próximas do início do estro podem resultar em taxas de fertilização mais baixas, mas sobrevivência embrionária mais alta. Juntos, estes fatos sugerem que é importante alguma seleção natural para a fertilização pelo espermatozóide "mais apto". Há relatos de incidências mais altas de polispermia e aumento do número de cromossomos após a inseminação de vacas superovuladas (King, 1985).
Já sabemos há anos que a mestiçagem acentua virtualmente todos os aspectos da reprodução. Isto se deve ao valor genético aditivo e não aditivo que resulta da mestiçagem. Quando cruzamos duas raças, o valor genético aditivo destes cruzamentos resulta em uma progênie que é a média dos pais. Este valor genético aditivo é tudo o que obtemos com a mestiçagem para algumas características (como as características de carcaça). Contudo, o valor genético não aditivo destes cruzamentos resulta em melhorias na progênie, que é melhor do que a média dos pais (as características de produção são exemplos). Algumas vezes, o valor genético não aditivo destes cruzamentos resulta em melhoria tão dramática que a progênie é melhor em certas características do que qualquer um dos pais.
A reprodução é um dos exemplos em que o vigor híbrido da mestiçagem resulta em melhora no desempenho reprodutivo. É provável que nem toda esta melhora esteja relacionada com uma melhora na sobrevivência embrionária, mas pelo menos uma parte pode ter relação. As anomalias genéticas também podem causar mortalidade embrionária tardia (dia 27 a 42) ou mortalidade fetal (depois do dia 42 da gestação). A mortalidade fetal estava aumentada entre bovinos Hereford quando se comparou animais endogâmicos com os não endogâmicos (MacNeil et al., 1989). Além disso, com o aumento do nível de endogamia também aumentou o nível de mortalidade fetal.
O estágio exato da gestação em que ocorreu a mortalidade fetal no estudo acima não é conhecido. Cundiff et al. (1974) relataram que a safra de bezerros foi 6,4% maior para as vacas mestiças do que as vacas puras, sendo que esta vantagem resulta de diferenças nas taxas de prenhez e de concepção ao primeiro serviço. As duas linhas de evidência sugerem que a mestiçagem deveria resultar em aumento da sobrevivência embrionária e fetal e provavelmente isto ocorre.










